A special thanks to The Washington University School of Medicine
SPINAL MOTOR STRUCTURES
A. The spinal cord:
Although we usually study the spinal cord as a
series of cross sections, it is important to remember that it is in fact a
column, with continuous tracts and cell columns. However, the cord can be
divided into segments by the nerve roots that come off of it; although the
rootlets branch off nearly continuously, they coalesce into about 31 discrete
nerves along the cord (8 cervical, 12 thoracic, 5 lumbar, 5 sacral, and 1
coccygeal nerves). At each segment, rootlets appear to come out of both the
dorsal and ventral halves of the spinal cord, as you see here:
In fact, only the ventral roots are coming out of the cord - the dorsal roots are actually going in. Throughout the cord, the dorsal grey matter (dorsal horns) deals with sensory perception, and receives information from the periphery through the dorsal root. The ventral horns contain the a-motor neurons, whose axons exit the cord via the ventral roots and travel directly to the muscles.
Along the dorsal root is a collection of cell
bodies called the dorsal root ganglion. Inside the ganglion are the cell
bodies of all the receptor neurons that send processes out to the periphery. The
free nerve ending in the tip of your finger that feels the paper cut actually
has its cell body back in the dorsal root ganglion. As you can see from the
picture, the dorsal root ganglion is actually located ventrally, but you can
tell that it is part of the dorsal root.
B. Levels of the spinal cord:
By this time in the course you have probably
noticed that different levels of the cord are different in shape. Could you
identify the source of a section if you had nothing to compare it to? In
general, you should be able to differentiate cervical from thoracic from lumbar
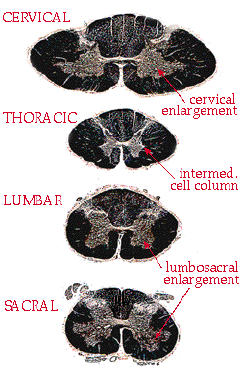
from sacral. Here is a series of cross sections:
|
The first thing to notice is
overall shape. Cervical sections tend to be wide and squashed
looking, like an oval. Compare the cervical section to the round lumbar
section.
The second thing to check for is a ventral horn enlargement. At segments that control a limb, the motor neurons are large and numerous. This causes enlarged ventral horns in two places: the lower cervical sections (C5-C8) and the lumbar/sacral sections. If you see an enlargement, you just need to differentiate cervical from lumbar. This can be done by shape (see above) or by proportion of white matter. The amount of white matter relative to grey matter decreases as you move down the cord. This is logical - in the white matter of the cervical cord you have all of the axons going to or from the entire body, more or less. In sacral cord the white matter contains only |
| those axons going
to or from the last couple of dermatomes - all other axons have "gotten off"
at higher levels. This is why sacral cord looks like it has so much grey
matter - really it has just lost all of the white. So, in summary, here are the level cues so far: wide flat cord, lots of white matter, ventral horn enlargements = cervical. Round cord, ventral horn enlargements = lumbar. Small round cord, almost no white matter = sacral. And the remaining level, thoracic, is the easiest of all. Notice the pointed tips which stick out between the small dorsal and ventral horns. This extra cell column is called the intermediate horn, or the intermediolateral cell column. It is the source of all of the sympathetics in the body, and occurs only in thoracic sections. |
|
C. Muscle spindles and the myotatic reflex:
One of the most familiar reflexes is the stretch reflex, also known as the knee-jerk reflex and the myotatic reflex. In its simplest form it is a 2-neuron loop, one afferent neuron and one efferent neuron. The afferent neuron is connected to a muscle spindle, which detects stretch in the muscle. The efferent neuron is the motor neuron, which causes the muscle to twitch.

But there are actually several types of afferents reporting on the status of the muscle. Let's look more closely at the muscle spindle:
The muscle spindle is a small group of muscle
fibers walled off from the rest of the muscle by a collagen sheath. The sheath
has a spindle or "fusiform" shape, so these fibers are called intrafusal
fibers, and are contrasted with the extrafusal fibers, which are the
power-generating muscle fibers. There are two types of nerve endings wrapped
around this intrafusal fiber, both of which monitor its degree of stretch - as
the muscle stretches, so does this capsule within it. These two stretch
receptors are sensitive to different time scales, however. The first is
called a Ia (that's one-A) fiber; the classification scheme is based on
diameter and conduction velocity, so the Ia's are the largest and fastest. The
Ia fiber fires like crazy when the muscle is stretching, but it is rapidly
adapting. As soon as the muscle stops changing length, and holds a new
position, the Ia adapts to the new length and stops firing. But you also need to
know the position of your muscle when it is still. The second type of stretch
receptor is called a II fiber, and it is slowly adapting. It also responds when
the muscle is stretching, but it maintains a firing rate after the muscle has
stopped moving (essentially, it is non-adapting). This information is part of
what allows you to tell the position of your arm when your eyes are closed.
Rule of nomenclature - if there is a Ia, where
is Ib? The Ib fibers are not connected to muscle spindles at all. Instead
they are embedded in the tendon, and monitor overall muscle tension from there.
They are also called Golgi tendon organs (the word "Golgi" is littered
throughout neuroanatomy - he was a famous early anatomist).
Now, there is a potential problem with the
muscle spindle system which may have occurred to you. What happens when the
muscle gets shorter? Does the spindle go limp and slack? How can it remain
sensitive to stretch at short lengths? This is where the intrafusal fiber comes
into play. Like any muscle fiber, it can contract. When it contracts, the entire
spindle shortens, remaining taut, and the sensitivity is intact. There are small
motor neurons in the ventral horn that innervate the intrafusal muscle fibers
and cause them to contract - they are the
g-motor
neurons. These neurons are excited
every time the a-motor
neurons fire, so that as the muscle contracts, the intrafusals contract with it.
How are they all hooked together? There are two simple rules: 1) When the stretch receptors fire, the a-motor neuron is excited, and the muscle contracts.
2) When the Golgi tendon organ fires, the a-motor neuron is inhibited (via an inhibitory interneuron), and the muscle relaxes. The purpose here is that the stretch receptors tell the muscle when it needs a little more force - that despite intending to contract the muscle is lengthening. This helps you to maintain the correct muscle tone. The Golgi tendon organs, on the other hand, begin to fire when the tension on the tendon is so great that you are in danger of injury. They have a protective function, and therefore they tell the muscle to ease off before it tears.

Occasionally, especially in cases of pyramidal
tract damage, these two systems can get stuck in a loop, where they alternately
trigger each other, causing the muscle to contract-relax-contract-relax, several
times a second. This rapid trembling is called clonus, and can be a sign
of pathology or extreme muscle fatigue.
D. Multiple motor pathways in the cord:
There are several pathways which innervate the a-motor neurons. They can be roughly grouped into the voluntary motion pathways and the postural pathways. The voluntary pathways include the lateral and anterior corticospinal systems, as covered in the "Basic motor" section. The postural pathways do not originate in cortex; instead their function is to maintain an upright posture against gravity, a task which requires hundreds of little muscular adjustments that we are not aware of. There are three principal pathways in humans: the vestibulospinal, tectospinal, and reticulospinal pathways. Pathways are always named beginning-to-end, so these originate in the vestibular nuclei, tectum (superior colliculi), and reticular formation, respectively. The rubrospinal system (from the red nucleus) is also sometimes included, but in humans it may be insignificant.
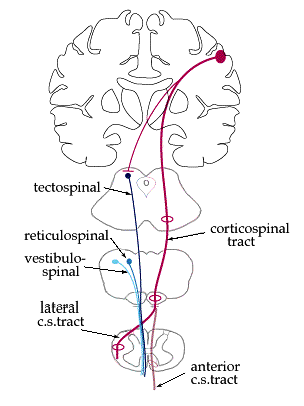
Although these pathways do not originate in
cortex, they are controlled to some degree by cortical structures. You must be
able to turn off selective postural systems to accomplish other movements. This
becomes apparent when the cortex is damaged or cut off from the postural
pathways, and can no longer control them. If the cortical input is damaged close
to its source, (i.e., in the internal capsule), the result is what is called a
decorticate posture. Here the postural pathways flex the upper limbs and
extend the lower limbs by default, since they are getting no input from cortex.
If the damage cuts off not just cortical input
but all input from the entire cerebrum, such as with a massive brainstem injury,
the result is a decerebrate posture. In the decerebrate position all four
limbs are extended and somewhat turned in (pronated). This sort of injury is
much more serious than an injury of the internal capsule; the prognosis is
usually very poor.
E. Injury to the corticospinal tract:
Just as the postural pathways have a "mind of
their own" when cortical control is cut off, the spine can also produce some
weird behaviors when the corticospinal tract is damaged. All of the spinal
reflexes are local - all of the cells involved are contained within one or two
segments, and cortex is not necessary. Therefore reflexes would still be present
in a transected spinal cord. However, the cortex normally keeps a tight rein on
reflexive behavior, so that it doesn't interfere with normal movements. When the
cortex is cut off, the spinal cord becomes hyperreflexic. All of the
normal reflexes become exaggerated, and some new ones appear. For example,
stroking the lateral sole of your foot with a sharp object would normally make
your toes curl downward. In a patient with corticospinal damage (also called
upper motor neuron damage), the big toe would lift up and the toes would fan
out. This is called the Babinski sign, and it is always pathological
(with the exception of very young infants).