A special thanks to The Washington University School of Medicine
BASAL GANGLIA
The basal ganglia and cerebellum are large collections of nuclei that modify movement on a minute-to-minute basis. Motor cortex sends information to both, and both structures send information right back to cortex via the thalamus. (Remember, to get to cortex you must go through thalamus.) The output of the cerebellum is excitatory, while the basal ganglia are inhibitory. The balance between these two systems allows for smooth, coordinated movement, and a disturbance in either system will show up as movement disorders.
A. The basal ganglia:
What are the basal ganglia? The name is
confusing, as generally a ganglion is a collection of cell bodies outside
the central nervous system. Blame the early anatomists. The basal ganglia are a
collection of nuclei deep to the white matter of cerebral cortex. The name
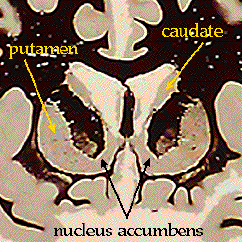
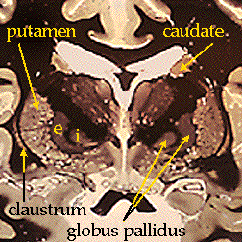
includes: caudate, putamen, nucleus accumbens, globus pallidus, substantia
nigra, subthalamic nucleus, and historically the claustrum and the
amygdala. However, the claustrum and the amygdala do not really deal with
movement, nor are they interconnected with the rest of the basal ganglia, so
they have been dropped from this section. Other groupings you may hear are the
striatum (caudate + putamen + nucleus accumbens), the corpus striatum (striatum
+ globus pallidus), or the lenticular nucleus (putamen + globus pallidus), but
these groupings obviously get confusing very quickly, so we will try to avoid
them.
The anatomy of these structures should be a
review from the "coronal and horizontal sections" lab. Here once again are the
basal ganglia as they appear when stained for myelin:
rostral section: |
middle section: |
|
|
caudal section:
|
|
An alternate stain is the acetylcholinesterase (AChE) stain. This technique stains for the enzyme that degrades acetylcholine (ACh), a major neurotransmitter. Areas which use ACh generally stain darkly. Here is a section through monkey brain, stained for AChE.
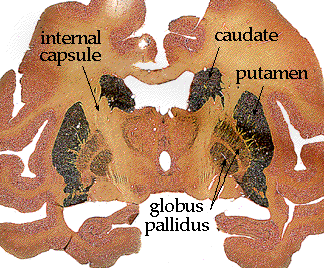
You can see that the caudate and putamen are
stained, while the globus pallidus remains fairly pale. This emphasizes their
different functions and connections. And those are...?
B. Different functions and connections:
The relationships between the nuclei of the
basal ganglia are by no means completely understood. When dealing with the
brain, you may sometimes be tempted to think that everything is connected to
everything else. Take heart, some fairly simple generalizations and schematics
can be drawn.
The caudate and putamen receive most of the
input from cerebral cortex; in this sense they are the doorway into the basal
ganglia. There are some regional differences: for example, medial caudate
and nucleus accumbens receive their input from frontal cortex and limbic areas,
and are implicated more in thinking and schizophrenia than in moving and motion
disorders. The caudate and putamen are reciprocally interconnected with the
substantia nigra, but send most of their output to the globus pallidus (see
diagram below).
The substantia nigra can be divided into two
parts: the substantia nigra pars compacta (SNpc) and the substantia
nigra pars reticulata (SNpr). The SNpc receives input from the caudate and
putamen, and sends information right back. The SNpr also receives input from the
caudate and putamen, but sends it outside the basal ganglia to control head and
eye movements. The SNpc is the more famous of the two, as it produces dopamine,
which is critical for normal movement. The SNpc degenerates in Parkinson's
disease, but the condition can be treated by giving oral dopamine precursors.
The globus pallidus can also be divided into
two parts: the globus pallidus externa (GPe) and the globus pallidus
interna (GPi). Both receive input from the caudate and putamen, and both are
in communication with the subthalamic nucleus. It is the GPi, however, that
sends the major inhibitory output from the basal ganglia back to thalamus. The
GPi also sends a few projections to an area of midbrain (the PPPA), presumably
to assist in postural control.
This schematic summarizes the connections of
the basal ganglia as described above.

Although there are many different
neurotransmitters used within the basal ganglia (principally ACh, GABA, and
dopamine), the overall effect on thalamus is inhibitory. The function of the
basal ganglia is often described in terms of a "brake hypothesis". To sit still,
you must put the brakes on all movements except those reflexes that maintain an
upright posture. To move, you must apply a brake to some postural reflexes, and
release the brake on voluntary movement. In such a complicated system, it is
apparent that small disturbances can throw the whole system out of whack, often
in unpredictable ways. The deficits tend to fall into one of two categories: the
presence of extraneous unwanted movements or an absence or difficulty with
intended movements.
C. Lesions of the basal ganglia:
Lesions in specific nuclei tend to produce
characteristic deficits. One well-known disorder is Parkinson's disease,
which is the slow and steady loss of dopaminergic neurons in SNpc. An instant
Parkinson-like syndrome will result if these neurons are damaged. This happened
several years ago to an unfortunate group of people who took some home-brewed
Demerol in search of a high. It was contaminated by a very nasty byproduct, MPTP
,which selectively zapped the SNpc neurons. The three symptoms usually
associated with Parkinson's are tremor, rigidity, and
bradykinesia. The tremor is most apparent at rest. Rigidity is a result of
simultaneous contraction of flexors and extensors, which tends to lock up the
limbs. Bradykinesia, or "slow movement", is a difficulty initiating voluntary
movement, as though the brake cannot be released.
Huntington's disease, or chorea, is a hereditary disease of unwanted movements. It results from degeneration of the caudate and putamen, and produces continuous dance-like movements of the face and limbs. A related disorder is hemiballismus, flailing movements of one arm and leg, which is caused by damage (i.e., stroke) of the subthalamic nucleus.
As with brain research generally, much of what is known about the basal ganglia comes from studying diseases which affect them. This section briefly outlines the range of diseases which are caused by damage to the basal ganglia.
Damage to both the basal ganglia and the supplementary motor area are correlated with impaired performance on sequential tasks [6].
The ``theme'' of diseases affecting the basal ganglia appears to be that the balance between the two major pathways is disturbed: the result is either involuntary movements or impairments to motion.
According to Kandel et al. [32], the impaired motions include ``lack of movement (akinesia), slowness of movement (bradykinesia), and the shuffling gait of Parkinson's disease.''
The list of involuntary movements includes
tremors (rhythmic, involuntary, oscillatory movements), athetosis (slow, writhing movements of the fingers and hands, and sometimes of the toes), chorea (abrupt movements of the limbs and facial muscles), ballism (violent, flailing movements), and dystonia (a persistent posture of a body part which can result in grotesque movements and distorted positions of the body).
The major diseases of the basal ganglia are [32]: