A special thanks to The Washington University School of Medicine
BASIC MOTOR PATHWAY
A. An overview:
The motor pathways are pathways which
originate in the brain or brainstem and descend down the spinal cord to control
the a-motor
neurons. These large neurons in the
ventral horns of the spinal cord send their axons out via the spinal roots and
directly control the muscles. The motor pathways can control posture, reflexes,
and muscle tone, as well as the conscious voluntary movements that we think of
when we hear "motor system". The most famous pathway is the so called "pyramidal
system", which begins with the large pyramidal neurons of the motor cortex,
travels through the pyramids of the brainstem, (somewhere in here there is a
coincidence), and finally ends on or near the
a-motor
neurons. This system is extremely important clinically, as strokes often affect
the motor system. Therefore it is crucial to understand the anatomy of the motor
pathway.
B. The anatomy:
The primary motor pathway is also called the
corticospinal pathway. As all such pathways are named from beginning to
end, this pathway starts in cortex and ends in the spine. Specifically, it
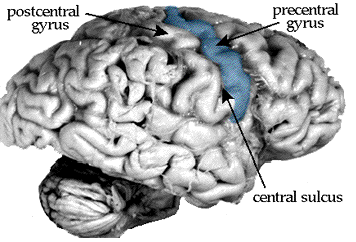
starts in the precentral gyrus, the fold of cortex just anterior to the
central sulcus.
The precentral gyrus has many names: primary
motor cortex, Brodmann's area 4, M1, etc. It provides the bulk of the
corticospinal tract, but other cortical areas contribute as well. One such area
is area 3a, part of primary somatosensory cortex, which is hidden down
inside the central sulcus.
If we take a section through the central
sulcus, we can see subtle differences between the pre- and post-central gyri.
All areas of cerebral cortex have six varying layers of cells, from the most
superficial and cell-free layer I to the deep layer VI. Each layer has a
slightly different cellular makeup, and the thicknesses of the layers vary with
cortical area. The postcentral gyrus, also known as primary somatosensory
cortex, has a distinct layer IV - across cortical areas, layer IV generally
receives sensory information. In the precentral gyrus, the motor cortex doesn't
receive much in the way of sensory input, so layer IV is indistinct. However,
layer V, which is generally responsible for sending information down to the
brainstem and beyond, is very prominent in motor cortex.
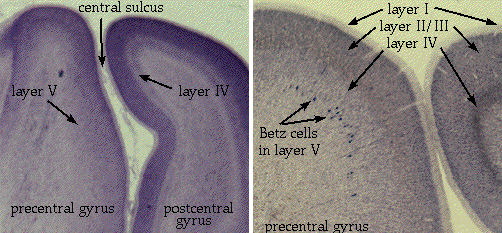
The corticospinal tract originates as the
axons of pyramidal neurons in layer V of (mainly) primary motor cortex. In
general, the farther the axon has to go, the larger the neuron. In the
precentral gyrus you can see some especially large neurons, visible even at low
magnification. These neurons are called Betz cells, and were once thought
to be the sole source of the corticospinal tract. We now know that they are only
a subset of the pyramidal neurons which make up the tract. Pyramidal neurons,
incidentally, occur in all types of cortex - it is just a morphological name for
large neurons that are triangular in shape, pointing a long apical dendrite
toward the surface of the brain.
Once the axons leave the pyramidal cells, they enter the white matter just below layer VI. Every gyrus in the brain has this core of white matter, which contains all of the axons entering or exiting the gyrus. As you get deeper into the brain, all of these slips of white matter coalesce to form one large body of axons, the corona radiata, "radiating crown". As you get still deeper into the hemispheres, the corona radiata dives into the deep nuclei of the brain, the caudate and putamen, splitting them in two. At this point, all of these axons are called the internal capsule.
The internal capsule is a major two-way
highway, and very vulnerable to strokes. Sensory information travels up it on
the way from the thalamus to the cortex, and motor information travels through
on the way down to the spine. As you saw in the horizontal sections at the
beginning of the course, the internal capsule has an anterior and posterior
limb. The motor and somatosensory information travels through the posterior
limb.
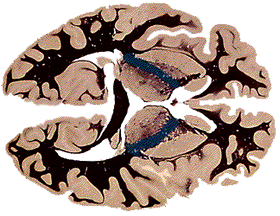
If you were to follow horizontal sections down
through the brain, at around the level where the midbrain begins, you
would see the internal capsule coalesce into a tight bundle to exit the cerebral
hemispheres. At this point the axons are called the cerebral peduncles,
or the "stalks" of the cerebrum. The peduncles make up the floor of the
midbrain, and contain all of the descending axons going to the brainstem or
spine. The peduncles, unlike the internal capsule, are largely one-way; most of
the axons in them are heading south. The ascending, sensory axons take other
routes to get to the thalamus.
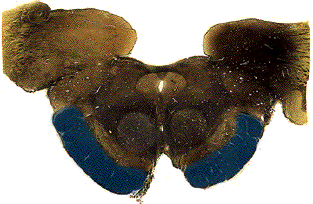
Once midbrain gives way to pons, two things happen to the peduncles. One, many of the axons from cortex were actually headed for the pons (the "corticopontine" fibers, of course), so they get off and synapse. Two, the remaining corticospinal axons get a little fragmented in the pons, so they are no longer visible as a nice tight bundle. They can be seen as several smaller bundles, though.
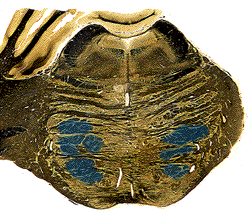
In the medulla, the fibers come together again as the pyramids. The pyramids were actually named as landmarks on the surface of the brainstem - on a human brainstem you can clearly see them as two ridges running down the ventral midline. The pyramids run the entire length of the medulla, large uninterrupted axon tracts on the ventral surface.
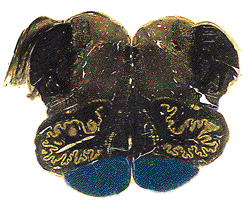
At the very caudal-most end of the medulla, right about at the point where you have to start calling it cervical spinal cord, the fibers in the pyramids cross. The cerebrum controls the opposite side of the body, so it had to happen sometime, didn't it? The crossing event is called the decussation of the pyramids, and you can identify it by the way the midline groove is suddenly way off the midline. In any single section, it looks like only one side is crossing, but this is a sectioning artifact. Large bundles of axons take turns crossing, much like your fingers interlace when you clasp your hands. As you section down through the decussation, the midline fissure will bend first to one side, then the other.

As each individual fiber crosses (remember, these are still the same axons we started with, from the pyramidal neurons of area 4!), where does it go? The pyramids do not merely exchange places. Instead each crossing axon takes up residence in the lateral white matter of the spinal cord. By the time the decussation is completed, the corticospinal fibers reside in this new location, now called the lateral corticospinal tract.
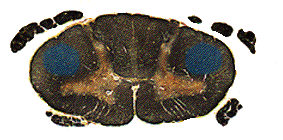
From this position they dive into the grey
matter of the spinal cord at their target levels. Those fibers controlling the
arms, for example, get off in the cervical levels of the cord. Once in the
ventral horn they synapse either on interneurons (most common) or directly
on the a-motor
neurons. They preferentially innervate the limbs and distal muscles.
C. A lesson in nomenclature:
As you may have learned in anatomy, if there is a greater horn of Ubu, there is probably also a lesser horn of Ubu. (Fictional, don't look it up.) Likewise, if there is a lateral corticospinal tract, there must also be a medial one. Indeed there is, only it is called the anterior corticospinal tract. Take comfort in the fact that it does actually lie medially. These fibers were part of the original corticospinal tract, and made up 15-20% of the pyramids, but at the decussation, they did not cross. As a result, in the spinal cord they are still sitting right where the pyramids were - ventrally, on either side of the midline.
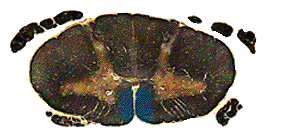
These fibers also dive into the ventral horns
at appropriate levels, and they tend to innervate the muscles of the trunk. Fine
finger movements and tap dancing are the exclusive domain of the lateral
corticospinal tract, but push-ups and hula-hooping rely more on the anterior
corticospinal tracts.
D. In summary:
Here is a diagram summarizing the two main
pathways of the voluntary motor system:
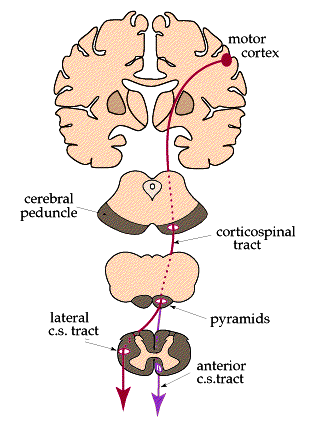
E. Clinical notes:
In motor cortex, the body is mapped out across the extent of the gyrus. Control of the feet lies near the midline at the top of the gyrus, whereas the lateral side of the gyrus controls the hands and face. Because the body parts are spread so widely in cortex, cortical strokes are rarely big enough to affect an entire half of the body. Instead, the hands or face, which take up the most cortical area, are preferentially affected. Strokes in the internal capsule are a different story. Fibers are bundled together so closely that a small stroke can in fact paralyze an entire side of the body - this is called hemiplegia (half-paralysis) or hemiparesis (half-weakness). If a stroke occurs in the brainstem, affecting the pyramids, the hemiparesis will often be accompanied by other symptoms or "neighborhood signs"; the brainstem is a very crowded place.
Although the cortex is primarily concerned
with the contralateral (opposite) side of the body, some muscle groups get
innervation from both hemispheres. This bilateral innervation protects the
muscle groups in case of stroke. Most of the muscles in the trunk are
bilaterally innervated, so that even hemiplegic patients can hold their torsos
upright. For some reason, the forehead also receives bilateral innervation,
which can be a key clue in determining if a patient's facial paralysis is due to
a cerebral stroke or a peripheral nerve injury. In a cerebral stroke patient,
only the lower half of the face (on one side) will be affected. In a patient
with Bell's palsy (a temporary inflammation of the facial nerve), the entire
side of the face will be affected.